Det gjør at avlsverdiene ikke kan identifiseres direkte, men beregnes ved hjelp av en statistisk modell. En slik modell fungerer både for kvantitative og kvalitative egenskaper, og brukes derfor for å beregne avlsverdier for alle egenskaper.
Den sanne avlsverdien
Den sanne avlsverdien til et dyr for en bestemt egenskap er summen av de additivt virkende genene til dyret som påvirker denne egenskapen. I beregningene av avlsverdier prøver man å komme så nærme den sanne avlsverdien som mulig. Sikkerheten på en avlsverdi sier noe om hvor nær den beregnede avlsverdien er den sanne avlsverdien. Den sanne avlsverdien kommer man aldri til å vite helt eksakt hva er.
For å komme så nærme den sanne avlsverdien som mulig når vi beregner avlsverdier, kreves det registrering og lagring av alle relevante data på individnivå, identifikasjon av dyret selv og alle foreldredyr flere generasjoner tilbake, samt passende statistiske modeller og kraftige datamaskiner.

Alt som kan observeres direkte på dyret kaller vi fenotype. Dette er en kombinasjon av arv og miljø (genotypen til dyret og miljøet rundt dyret). Foto: Els Korsten.
Hvilke gener som uttrykkes, påvirker fenotypen
Den additive genetiske effekten er den arvelige delen av genene. Men hvilke gener som kommer til uttrykk kan påvirkes av dominans og epistasi. Man arver alltid dobbelt sett med gener, et sett fra far og et sett fra mor.
Dominans vil si at et gen dominerer over et annet gen. Det betyr at to dyr, hvor det ene er homozygot (har to like genvarianter) og det andre er heterozygot (har to ulike genvarianter) for et bestemt gen, prestere like godt i samme miljø for en egenskap, men genetisk vil dyrene være ulike.
Epistasi vil si at et gen blir påvirket av et eller flere andre gen (genetisk bakgrunn). Dette kan gjøre at et slik gen kan komme til uttrykk på ulike måter, om ikke den genetiske bakgrunnen er lik hos alle individer.
Når man krysser inn avlsmateriale fra andre populasjoner vil med stor sannsynlighet den beste genvarianten komme til uttrykk. Derfor kan avkommet yte bedre enn det den additive genetiske effekten (avlsverdien) skulle tilsi. Summen av dominanseffektene for hvert gen kalles heterosis eller krysningsfrodighet.
Effekt av miljø
Alle kjente miljøfaktorer som påvirker fenotypene må registreres og korrigeres for. Hvis ikke kan effekten av miljøfaktorene bli tatt for å være en del av de genetiske forskjellene mellom grupper av dyr, noe som kan føre til at avlsverdiene blir overestimert eller underestimert. I statisiske modeller blir miljøeffekter skilt i ulike grupper, avhengig av definisjonen på effektene (eksempelvis faste og tilfeldige miljøeffekter).
Et eksempel på en miljøeffekt er laktasjonsnummer. Hvis ei ku har hatt tre laktasjoner, og det er gjort gjentatte observasjoner på kua sin avdrått i hver laktasjon, må det korrigeres for miljøeffekter som har påvirket de gjentatte observasjonene. Hadde dette vært ei ku som ble trespent i første laktasjon, ville det dermed påvirke avdråtten i de neste laktasjonene negativt om det ikke hadde blitt tatt hensyn til i den statistiske modellen som beregner avlsverdier.
Estimert avlsverdi (beregnet avlsverdi)
En statistisk modell for en egenskap er i hovedtrekk bygget opp av fenotyper, miljøeffekter og slektskap mellom individer. Basert på denne informasjon blir det beregnet en estimert avlsverdi.
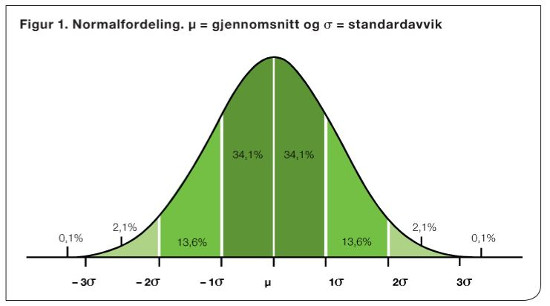
I modellen blir egenskapene definert som normalfordelte (Figur 1), som er en klokkeformet fordeling. Denne «klokkeformede» fordelingen er kjennetegnet ved at de fleste fenotypene vil være nær gjennomsnittet, og så vil det bli færre og færre av fenotypene desto lenger ut mot hver av endene man kommer.
Størrelsen på variasjonen til en normalfordeling uttrykkes ofte i standardavvik (kvadratroten av variasjonen). For hvilken som helst normalfordeling, uavhengig av art og egenskap, vil et standardavvik på hver side av gjennomsnittet tilsammen omfatte ca. 68 % av verdiene, to standardavvik vil omfatte ca. 95 % av verdiene og tre standardavvik 99% (Figur 1). For eksempel hvis det fenotypiske gjennomsnittet for melkemengde er 7 000 kg og standardavviket er 1 000 kg, så vil 68 % av fenotypene være mellom 6 000 og 8 000 kg og 99 % være mellom 4 000 og 10 000 kg.
Arvegrad
Arvegraden er et mål på hvor stor del av den genetiske variasjonen som forklarer den observerte fenotypiske variasjonen mellom dyr i en populasjon, og oppgis som en ratio mellom 0 og 1. Der 0 er ikke arvelig og 1 er fullstendig arvelig.
Når arvegraden er høy, vil den genetiske variasjonen utgjøre en stor andel av den fenotypiske variasjonen som observeres, og det vil være lettere å indentifisere gener som påvirker egenskapen. Man vil derfor raskere oppnå en sikrere avlsverdi for disse egenskapene sammenlignet med lavarvelige egenskaper. Lavarvelige egenskaper er avhengig av flere fenotypiske observasjoner for å få en sikker avlsverdi. For egenskaper med lav arvegrad spiller miljøet en stor rolle, og det kan være vanskeligere å skille mellom hva som skyldes genetikk og hva som skyldes miljø. Har man derimot mye informasjon (både fenotyper og genotyper) på dyret selv og slektninger vil man få beregnet avlsverdier og klare å skille de beste dyra også for disse egenskapene.
Genetiske sammenhenger (genetisk korrelasjon)
En genetisk korrelasjon angir graden av arvelig sammenheng mellom egenskaper. En korrelasjon er et tall mellom -1 og +1, der 1 betyr at det er samme egenskap og 0 betyr at det er ingen sammenheng. Hvis økning i den ene egenskapen har sammenheng med reduksjon av den andre egenskapen er korrelasjonen negativ. Omvendt, hvis sammenhengen gir økning eller reduksjon på begge egenskapene, er korrelasjonen positiv. Genetisk korrelasjon mellom to egenskaper skyldes et eller flere gen som påvirker begge egenskapene, eller at gener som påvirker flere egenskaper ligger nær hverandre og derfor nedarves sammen.
Seleksjon for en bestemt egenskap vil påvirke utvikling også for genetisk korrelerte egenskaper. Det vil si at man har en indirekte seleksjonsrespons for alle de genetisk korrelerte egenskapene. Dette kan være en gunstig eller ugunstig genetisk korrelasjon.
Gunstig genetisk korrelasjon betyr at avlsmessig utvikling på en egenskap påvirker en annen egenskap i ønsket retning. Ugunstig genetisk korrelasjon betyr at å avle på en egenskap gir ugunstig utvikling av en annen egenskap. En gunstig genetisk korrelasjon og en ugunstig genetisk korrelasjon kan begge ha både et positiv eller negativt fortegn. For eksempel er den genetiske korrelasjonen mellom kg melk og mastitt positiv, men ugunstig. Avler man for økt kg melk, avler man indirekte for flere mastitt-tilfeller, hvis ikke mastitt også tas hensyn til i avlsarbeidet.
Indekser og samla avlsverdi
En indeks er en standardisert avlsverdi, som er presentert som et avvik fra gjennomsnittet i populasjonen eller en bestemt referansepopulasjon. Vanligvis blir gjennomsnittlig indeks satt til 100, med et standardavvik på ±10 poeng. Nesten alle dyra vil få indekser mellom 70 og 130, som er innenfor 3 standardavvik fra gjennomsnittet.
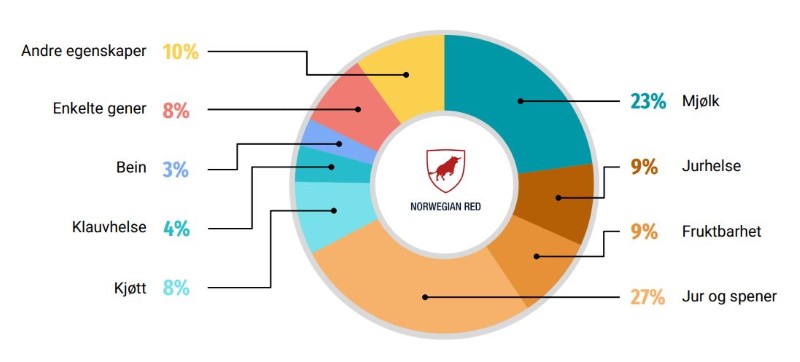
Egenskapene som vektes inn i samla avlsverdi.
Når et avlsmål skal bestemmes er det mange egenskaper som skal inngå. Egenskapene må derfor vektes inn i en samla avlsverdi, som skal gjenspeile ønsket respons hos hver av de enkelte egenskapene. Jo flere egenskaper som inngår i et avlsmål jo saktere vil egenskaper endre seg, fordi vektingene må fordeles på mange egenskaper. Omvendt, har man få egenskaper i et avlsmål, vil man oppnå ønsket respons på de enkelte egenskapene raskere fordi hver egenskap har større vektlegging. I denne sammenheng er det også viktig å ta hensyn til genetiske korrelasjoner mellom egenskaper. Egenskapene som vektes i avlsmålet vil gjennom genetiske korrelasjoner indirekte påvirke utviklingen av andre genetiske korrelerte egenskaper. Disse egenskapene kan både være andre egenskaper inkludert i avlsmålet eller det kan være egenskaper som ikke er vektlagt.


